Пресмыкающиеся
| Пресмыкающиеся | |||||||||||||||||||||
| |||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
промежуточные ранги
| |||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||
Reptilia Laurenti, 1768 | |||||||||||||||||||||
| Современные отряды | |||||||||||||||||||||
| |||||||||||||||||||||
| |||||||||||||||||||||




Пресмыка́ющиеся, или репти́лии (лат. Reptilia), — класс (по традиционной классификации) или парафилетическая группа (по кладистической классификации)[1] преимущественно наземных позвоночных животных, включающий современных черепах, крокодилов, клювоголовых, амфисбен, ящериц и змей. В XVIII—XIX веках вместе с амфибиями объединялись в группу гады — холоднокровные наземные позвоночные. Традиционно к этой группе относили различных позвоночных, по изначальным представлениям похожих по своей организации на современных рептилий (например, синапсидов — предков современных млекопитающих). Однако в настоящее время вопрос об физиологии многих вымерших крупных групп организмов остаются открытыми, а данные об эволюционных связях этих групп не поддерживают подобного рода классификацию. Многие авторы считают, что архозавров (крокодилов, птерозавров, динозавров и т. д.) следует вынести из класса пресмыкающихся и объединить их в один класс с птицами (которые с точки зрения кладистики в любом случае являются архозаврами)[2], или что птиц следует объединить с рептилиями в одну таксономическую группу (например, в класс Sauropsida)[3]. В мире известно около 9400 видов пресмыкающихся[4], на территории России обитает 77 видов[5][6].
Крупнейшие наземные животные принадлежали к динозаврам — представителям древних пресмыкающихся. Пресмыкающиеся пережили расцвет в эпоху мезозоя, когда они доминировали на суше, в море и в воздухе. В конце мелового периода большая часть пресмыкающихся вымерла. Современные рептилии — лишь разрозненные остатки того мира. Кроме того, они весьма примитивны и низкоорганизованны (за исключением крокодилов) по сравнению с многими вымершими видами. Однако, древние рептилии дали начало процветающей в настоящее время группе животных — птицам, причём многие приспособления, обусловившие эволюционный успех этой группы, появились ещё у её предков, представлявших собой специализированные формы рептилий (теплокровность, теплоизолирующий покров тела — перья, развитый головной мозг, и так далее).
Содержание
1 Строение
1.1 Покров
1.2 Скелет
1.3 Мышечная система
1.4 Нервная система
1.5 Органы чувств
1.6 Дыхательная система
1.7 Система кровообращения
1.8 Пищеварительная система
1.9 Выделительная система
1.10 Половая система
2 Образ жизни
2.1 Развитие
2.2 Питание
2.3 Передвижение
3 Голос
4 Хозяйственное значение
5 Эволюция пресмыкающихся
6 Филогенетика
7 Модельные объекты
8 Классификация
9 См. также
10 Примечания
11 Литература
12 Ссылки
Строение |
У большинства современных пресмыкающихся наблюдаются как черты более простых по строению амфибий, так и черты высших позвоночных животных.
Покров |

Чешуйки на коже ленточного крайта (Bungarus fasciatus)

Чешуйки на коже ящерицы.

Отпечатки перьев на фоссилии вымершей рептилии — микрораптора.
Наружный кожный покров современных пресмыкающихся в результате утолщения и ороговения образует чешуйки или щитки. У ящериц роговые чешуйки перекрывают друг друга, напоминая черепицу. У черепах сросшиеся щитки формируют сплошной прочный панцирь. Смена рогового покрова происходит путём полной или частичной линьки, которая у многих видов происходит несколько раз в год.
Наряду с этим, древние рептилии могли также быть покрыты своеобразными перьями (являющимися по строению специализированной формой чешуи) или похожими на пух нитевидными образованиями (прогрессивные архозавры).
Плотная и сухая кожа содержит пахучие железы. Слизистые железы отсутствуют — кожа рептилий сухая, характерный блеск ей придают гладкие чешуи.
В наружной части внутреннего слоя кожи часто находятся специальные клетки — хроматофоры. В этих клетках секретируются пигменты: меланины и каротиноиды. Также в хроматофорах встречается способный отражать свет гуанин. Благодаря хроматофорам, некоторые пресмыкающиеся способны изменять окраску своего тела за сравнительно короткое время. Хамелеоны — наиболее известные представители с подобным свойством.
Скелет |
В осевом скелете пресмыкающихся разделение на отделы более заметно, чем у земноводных. Хорошо различимы пять отделов позвоночника: шейный (лат. pars cervicalis), туловищный (пояснично-грудной, pars thoracolumbalis), поясничный, крестцовый (pars sacralis) и хвостовой (pars caudalis).
Типичным для пресмыкающихся является следующее строение осевого скелета. Общее количество позвонков различно у разных видов (50—80, у змей возрастает до 140—435). Из позвонков шейного отдела (от 7 до 10) два передних (атлант и эпистрофей) образуют сустав, позволяющий голове не только двигаться в вертикальной плоскости относительно первого шейного позвонка, но и поворачиваться. В туловищном отделе от 16 до 25 позвонков, каждый с парой рёбер. Первые несколько позвонков прикрепляются к грудине, образуя грудную клетку (отсутствует у змей). В крестцовом отделе всего два позвонка, к широким поперечным отросткам которых причленяется таз. Хвостовой отдел составляют несколько десятков (15—40) постепенно уменьшающихся в размерах позвонков. Последние хвостовые позвонки представляют собой небольшие палочковидные косточки.
В некоторых группах рептилий осевой скелет имеет отличия. У змей позвоночник отчетливо делится лишь на туловищный и хвостовой отделы, грудина отсутствует. У черепах позвонки туловищного отдела срастаются со спинным щитом панциря, вследствие чего неподвижны.
Череп пресмыкающихся значительно более окостеневший, чем у земноводных. Лишь в обонятельной капсуле и слуховой области содержится небольшое количество хряща. Осевой и висцеральный отделы черепа эмбрионально формируются отдельно, но у взрослых особей срастаются в единое образование. В состав черепа входят как хрящевые (замещающие, или первичные), так и многочисленные кожные (покровные, или вторичные) кости.
Пояс передних конечностей сходен с поясом земноводных, отличаясь лишь более сильным развитием окостенения. Пара передних конечностей рептилий состоит из плеча, предплечья и кисти. Пара задних конечностей — из бедра, голени и стопы. На фалангах конечностей расположены когти.
Мышечная система |
Мышечная система пресмыкающихся представлена жевательной, шейной мускулатурой, мускулатурой брюшного пресса, а также мускулатурой сгибателей и разгибателей. Присутствуют характерные для амниот межрёберные мышцы, играющие важную роль при акте дыхания. Подкожная мускулатура позволяет изменять положение роговых чешуй.
Энергоснабжение произвольной мускулатуры современных пресмыкающихся происходит главным образом за счёт анаэробного разложения глюкозы — порядка 50…75 % мышечных волокон в их мышцах приспособлены к работе в анаэробных условиях (без участия кислорода). Такой метаболизм позволяет им перемещаться на короткие расстояния столь же быстро, как и теплокровным животным, и обеспечивает даже большую силу сокращения мышц, причём интенсивность реакции у многих групп слабо зависит от температуры тела. Однако уже через 1-2 минуты интенсивной работы мускулатуры в анаэробном режиме в ней накапливается молочная кислота, что приводит к наступлению своеобразной мышечной усталости и изменению pH крови. В результате уже после нескольких быстрых пробежек рептилия становится практически неспособна к активным действиям на время от нескольких минут до нескольких часов, требующееся для распада накопившейся в мышцах молочной кислоты.[7] Более крупные животные способны дольше терпеть накопление в мышцах молочной кислоты, но в то же время не имеют преимущества в плане скорости восстановления мышц после физических нагрузок.
Некоторые современные рептилии, в частности, вараны и кожистая черепаха, тем не менее, являются исключением из этого правила: в ходе активности их уровень метаболизма может достигать более половины от такового у теплокровных животных сопоставимого веса, и они в значительной мере полагаются на аэробный метаболизм, почти не испытывая проблем с накоплением молочной кислоты.[8]
Среди вымерших древних рептилий известны высокоподвижные формы (такие как архозавры: наземные крокодиломорфы, динозавры и птерозавры, а также мозазавры, ихтиозавры и завроптеригии), строение тела и предполагаемый образ жизни которых говорят о способности поддерживать продолжительную мышечную активность без утомления, что соответствует более интенсивному, чем у современных пресмыкающихся, обмену веществ, и работе мышц преимущественно по аэробному механизму, требующему интенсивного снабжения кислородом. Это в свою очередь должно было сопровождаться совершенно другой физиологией организма (полное разделение желудочков сердца, повышение эффективности дыхания за счёт отделения воздухоносных путей от ротовой полости и появления вторичного нёба, появление эффективных механизмов терморегуляции и так далее). Данные изменения отчасти прослеживаются на палеонтологическом материале в линии рептилий, ведущей к птицам. В настоящее время надёжно показано существование теплокровности (эндотермной системы терморегуляции) как минимум у некоторых групп динозавров и предков современных крокодилов, а также плезиозавров и ихтиозавров.[9][7][10]
Нервная система |
Как и у большинства хордовых животных, центральная нервная система пресмыкающихся представлена головным (из 5 отделов) и спинным мозгом.
Головной мозг расположен внутри черепа. Ряд важных особенностей отличает головной мозг пресмыкающихся от головного мозга земноводных. Нередко говорят о так называемом завропсидном типе головного мозга, присущем также и птицам, в отличие от ихтиопсидного типа у рыб и земноводных.
Выделяют пять отделов головного мозга пресмыкающихся.
Передний мозг состоит из двух больших полушарий, от которых отходят обонятельные доли. Поверхность больших полушарий абсолютно гладкая. В мозговом своде полушарий различают первичный свод — архипаллиум, занимающий большую часть крыши полушарий, и зачатки неопаллиума. Дно переднего мозга в основном состоит из полосатых тел.
Промежуточный мозг расположен между передним и средним мозгом. В верхней его части расположен эпифиз, а на нижней стороне — гипофиз. У большинства ящериц и гаттерии (а также многих вымерших форм) рядом с эпифизом развивается теменной глаз, а у крокодилов потеряны оба этих органа. Дно промежуточного мозга занято зрительными нервами и их перекрестом (хиазмой).
Средний мозг представлен двумя крупными передними холмами — зрительными долями, а также небольшими задними холмами. Зрительная кора более развита, чем у амфибий.
Мозжечок прикрывает переднюю часть продолговатого мозга. Он более крупный по сравнению с мозжечком амфибий.
Продолговатый мозг образует изгиб в вертикальной плоскости, который характерен для всех амниот.
От головного мозга отходят 12 пар черепных нервов. В спинном мозге разделение на белое и серое вещество более отчётливо, чем у амфибий. От спинного мозга отходят сегментальные спинномозговые нервы, образуя типичное плечевое и тазовое сплетение. Отчётливо выражена вегетативная нервная система (симпатическая и парасимпатическая) в виде цепи парных нервных ганглиев.
Органы чувств |

Глаз геккона
Пресмыкающиеся имеют шесть основных органов чувств, из которых зрение имеет решающее значение.
Орган зрения — глаза, устроены сложнее, чем у амфибий: в склере присутствует кольцо из тонких костных пластинок; от задней стенки глазного яблока отходит вырост — гребешок, вдающийся в стекловидное тело; в ресничном теле развита поперечно-полосатая мускулатура, которая позволяет не только перемещать хрусталик, но и изменять его форму, таким образом осуществляя наводку на резкость в процессе аккомодации. Органы зрения имеют приспособления к работе в воздушной среде. Слёзные железы предохраняют глаза от высыхания. Наружные веки и мигательная перепонка выполняют защитную функцию. У змей и некоторых ящериц веки срастаются, формируя прозрачную оболочку. Сетчатка глаза может содержать как палочки, так и колбочки. У ночных видов колбочки отсутствуют. У большинства дневных видов диапазон цветного зрения смещён в жёлто-оранжевую часть спектра. Форма зрачка чаще всего круглая или в виде вертикальной щели (как у кошки), горизонтальный щелевидный зрачок, весьма распространённый у земноводных, среди рептилий встречается очень редко, например у плетевидных змей. Теменной глаз, сохранившийся у некоторых видов, важен для ориентирования в пространстве и синхронизации суточных ритмов организма с циклом смены дня и ночи, хотя многое в его функциях ещё неясно[11][12].
Орган обоняния представлен внутренними ноздрями — хоанами и вомероназальным органом. По сравнению со строением земноводных, хоаны расположены ближе к глотке, что даёт возможность свободно дышать в то время, как пища находится во рту. Обоняние развито лучше, чем у земноводных, позволяя многим ящерицам находить пищу, находящуюся под поверхностью песка на глубине до 6—8 см.
Орган вкуса — вкусовые луковицы, расположенные в основном в глотке.
Орган тепловой чувствительности находится на лицевой ямке между глазом и носом с каждой стороны головы. Особенно развит у змей. У ямкоголовых змей термолокаторы позволяют определять даже направление источника теплового излучения.
Орган слуха близок к органу слуха лягушек, он содержит внутреннее и среднее ухо, снабженное барабанной перепонкой, слуховой косточкой — стременем и евстахиевой трубой. Роль слуха в жизни пресмыкающихся сравнительно невелика, особенно слаб слух у змей, не имеющих барабанной перепонки и воспринимающих колебания, распространяющиеся по земле или в воде. Пресмыкающиеся воспринимают звуки в диапазоне 20—6000 Гц, хотя большинство хорошо слышит лишь в диапазоне 60—200 Гц (у крокодилов 100—3000 Гц).
Осязание выражено отчётливо, особенно у черепах, которые могут чувствовать даже лёгкое прикосновение к панцирю.
Дыхательная система |
Для пресмыкающихся характерно дыхание всасывающего типа путём расширения и сужения грудной клетки при помощи межрёберной и брюшной мускулатуры. Попавший через гортань воздух поступает в трахею — длинную дыхательную трубку, которая на конце делится на бронхи, ведущие в лёгкие. Как и у земноводных, лёгкие пресмыкающихся имеют мешкообразное строение, хотя их внутренняя структура намного сложнее. Внутренние стенки лёгочных мешков имеют складчатое ячеистое строение, что значительно увеличивает дыхательную поверхность.
Поскольку тело покрыто чешуйками, кожное дыхание у пресмыкающихся отсутствует (исключение составляют мягкотелые черепахи и морские змеи), и лёгкие являются единственным дыхательным органом.
Система кровообращения |
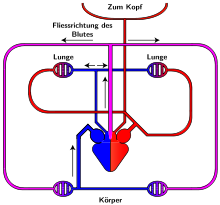
Система кровообращения пресмыкающихся
Как и амфибии, большинство пресмыкающихся обладают трёхкамерным сердцем, состоящим из желудочка и двух предсердий. Желудочек разделён неполной перегородкой на две половины: верхнюю и нижнюю. Однако, как известно трехкамерное сердце рептилий развилось независимо от такового у земноводных, и различается в плане строения (и иногда — функционала) у разных современных групп.[13][14]
При такой конструкции сердца в щелевидном пространстве вокруг неполной перегородки желудочка устанавливается градиент (разность) количества кислорода крови. После сокращения предсердий артериальная кровь из левого предсердия оказывается в верхней половине желудочка и вытесняет венозную кровь, излившуюся из правой части желудочка, в нижнюю половину. В правой части желудочка оказывается смешанная кровь. При сокращении желудочка каждая порция крови устремляется к ближайшему отверстию: артериальная кровь из верхней половины — в правую дугу аорты, венозная кровь из нижней половины — в лёгочную артерию, а смешанная кровь из правой части желудочка — в левую дугу аорты. Поскольку именно правая дуга аорты несёт кровь к мозгу, мозг получает наиболее обогащённую кислородом кровь.
Будучи архозаврами, крокодилы обладают четырёхкамерным сердцем, в котором лишь в ходе эмбриогенеза образуется отверстие Панница, допускающее контролируемое смешивание артериальной и венозной крови[10]. Перегородка полностью разделяет желудочек на две половины: правую — венозную и левую — артериальную, таким образом обосабливая полноценное четырёхкамерное сердце, как у млекопитающих и птиц.
В противоположность общему артериальному стволу земноводных, у рептилий наблюдается три самостоятельных сосуда: лёгочная артерия и правая и левая дуги аорты. Каждая дуга аорты загибается назад вокруг пищевода, и, сойдясь друг с другом, они соединяются в непарную спинную аорту. Спинная аорта тянется назад, отсылая по пути артерии ко всем органам. От правой дуги аорты, отходящей от левого артериального желудочка, ответвляются общим стволом правая и левая сонные артерии, от правой же дуги отходят и обе подключичные артерии, несущие кровь к передним конечностям.
Полного разделения на два независимых круга кровообращения у пресмыкающихся не происходит, поскольку венозная и артериальная кровь смешиваются в спинной аорте.
Подобно рыбам и амфибиям, подавляющее большинство современных пресмыкающихся— холоднокровные животные. Несмотря на это, многие пресмыкающиеся могут регулировать температуру, перемещаясь из тени на солнце и обратно или меняя цвет, становясь темнее, чтобы согреться, или светлее, чтобы остыть.
Сравнительно недавно у ящериц тегу была обнаружена факультативная эндотермия — возможность повышать температуру тела во время брачного периода за счет эндотермических химических реакций[15]. Вараны в разгар пищевой активности также могут повышать температуру тела за счет внутренних процессов[16]. Кожистая черепаха, в свою очередь, имеет изолирующую жировую прослойку и за счет интенсивной мышечной активности может быть примерно на 18 градусов теплее окружающей ее воды[17].
Пищеварительная система |
Вследствие разнообразия доступной для питания пищи пищеварительный тракт у пресмыкающихся гораздо более дифференцирован, чем у земноводных животных.
Пищеварительная система начинается ротовым отверстием, ограниченным челюстями с обычно коническими, одинаковыми, прочно прирастающими зубами (гомодонтная система, хотя у некоторых вымерших форм была гетеродонтная). Язык свободный, впереди мускулистый, подвижный, к концу утончается и раздваивается. Ротовая полость отграничена от глотки развивающимся вторичным костным нёбом у ящериц и черепах, полностью развитым костным небом у крокодилов, но у змей и варанов вторичное костное небо отсутствует. Многоклеточные слюнные железы содержат пищеварительные ферменты. Глотка переходит в узкий пищевод, далее в мускулистый желудок и кишечник. Желудок имеет толстые мышечные стенки. На границе между тонкой и толстой кишкой находится слепая кишка, которой нет у амфибий. Большая печень рептилий имеет желчный пузырь. Поджелудочная железа в виде длинного плотного тела лежит в петле двенадцатиперстной кишки. Кишечник заканчивается клоакой.
Выделительная система |
Выделительная система пресмыкающихся представлена почками, мочеточниками и мочевым пузырём.
Почки пресмыкающихся существенно отличаются от почек рыб и амфибий, которым приходится решать задачу избавления от постоянного избытка воды в организме. Вместо туловищных почек амфибий (мезонефрос), почки рептилий (метанефрос) располагаются в тазовой области с брюшной стороны клоаки и по её бокам. Почки соединяются с клоакой через мочеточники.
Тонкостенный стебельчатый мочевой пузырь соединяется с клоакой тонкой шейкой на её брюшной стороне. У некоторых пресмыкающихся мочевой пузырь недоразвит (крокодилы, змеи, некоторые ящерицы).
Также появляется новый орган выделения — тазовая почка.
У наземных рептилий конечный продукт азотистого обмена — мочевая кислота.
Половая система |
Пресмыкающиеся — раздельнополые животные, двуполое размножение.
Половая система самцов состоит из пары семенников, которые расположены по бокам поясничного отдела позвоночника. От каждого семенника отходит семенной канал, который впадает в вольфов канал. С появлением тазовой почки у пресмыкающихся вольфов канал у самцов выступает лишь как семяпровод и полностью отсутствует у самок. Вольфов канал открывается в клоаку, образуя семенной пузырёк.
Половая система самок представлена яичниками, которые подвешены на брыжейке к спинной стороне полости тела по бокам позвоночника. Яйцеводы (мюллеровы каналы) также подвешены на брыжейке. В переднюю часть полости тела яйцеводы открываются щелевидными отверстиями — воронками. Нижний конец яйцеводов открывается в нижний отдел клоаки на её спинной стороне.
Образ жизни |
Развитие |
Оплодотворение внутреннее. Развитие зародыша происходит в яйце с кожистой или известковой оболочкой, наряду с этим встречается яйцеживорождение и (реже) истинное живорождение. У пресмыкающихся прямое постэмбриональное развитие. Для многих представителей характерна забота о потомстве, в частности самки крокодилов переносят потомство от места кладки к водоёмам в ротовой полости, хотя в некоторых случаях могут съесть детёныша.
Питание |
Большинство пресмыкающихся относится к плотоядным животным (включая насекомоядность и поедание падали, истинное хищничество среди современных рептилий свойственно только крокодилам, ряду ящериц и большинству змей). Для некоторых (например, агамы, игуаны) характерно смешанное питание. Иногда встречаются и почти исключительно травоядные пресмыкающиеся (сухопутные черепахи).
Передвижение |

Протерозух, примитивный текодонт с конечностями, расположенными по бокам от туловища, как у современных рептилий, и приспособленными к «пресмыканию».

То же расположение конечностей у современной ящерицы прыткой.
Примитивные группы древних амниот (в том числе первые рептилии) изначально передвигались на четырёх широко расставленных в стороны конечностях, которые при локомоции раздвигались медиалатерально и не были способны постоянно поддерживать туловище над землёй, отсюда — «пресмыкание», волочение живота по земле, давшее русскоязычное название всей этой группе, что соответствовало характерному для них низкому уровню интенсивности обмена веществ и сравнительно малоподвижному образу жизни, связанному с чередованием коротких вспышек активности и длительного отдыха лёжа на животе. Современные пресмыкающиеся в основном сохраняют подобный, примитивный в эволюционном плане, характер локомоции, некоторое исключение составляют только крокодилы, у которых конечности при беге могут занимать положение почти под туловищем. Причем у некоторых вымерших групп современного отряда крокодилов, которые вели исключительно сухопутный образ жизни, такое положение конечностей было габитуальным, то есть поддерживалось большую часть времени — в Австралии такие наземные крокодилы (например, Квинкана) обитали ещё 40 тыс. лет назад, то есть одновременно с человеком современного вида.
Встречаются специализированные ползающие формы — змеи, безногие ящерицы. Большинство рептилий неплохо плавают за счёт «рыбообразного» изгибания тела в поперечной плоскости. У некоторых групп приспособление к водной среде достигло более высокой степени, причём в процессе эволюции у них вырабатывались различные типы локомоции. Так, ихтиозавры, мозазавры и, в меньшей степени, крокодилы сохранили характер плавания за счёт поперечных изгибов туловища. С другой стороны, у морских черепах и вымерших плезиозавров выработался новый механизм движения в воде, за счёт взмахов конечностей, преобразовавшихся в ласты. Многие эти группы практически полностью утратили способность перемещаться на суше. Несколько родов пресмыкающихся способны к планирующему полёту — например летучие драконы (агамы рода Draco) и древесные летучие змеи из рода Chrysopelea. Активно летающие рептилии известны только из ископаемых останков (см. Птерозавры).
В древности развитие некоторых продвинутых групп рептилий шло в иных направлениях — их конечности стали перемещаться непосредственно под туловище, занимая более габитуально вертикальное (выпрямленное) положение, что соответствовало более подвижному образу жизни, связанному с быстрым бегом и активным преследованием добычи.

Орнитозух, текодонт с конечностями, приспособленными к бегу на двух ногах.
У текодонтов лобковая и седалищная кости начали удлиняться, передние ноги — укорачиваться, а тяжёлый длинный хвост, практически лишившийся подвижности из-за срастания тазовых костей, превратился в балансир, уравновешивающий массу вытянутого туловища. Задние конечности при ходьбе стали перемещаться в парасагиттальной плоскости, голеностопный сустав получил возможность пружинить при ходьбе, что способствовало более экономному расходу сил, а опора переместилась на средний (третий) палец. Это позволило многим группам текодонтов перейти к факультативной, или даже габитуальной, бипедальности, подобно современным бегающим птицам, специализировавшись на беге на двух ногах.
Данные изменения должны были сопровождаться значительным повышением интенсивности обмена веществ, поскольку постоянное поддержание тела над землёй и длительная локомоторная активность требовали намного больших энергозатрат по сравнению с «пресмыканием». Однако проследить подобные изменения на палеонтологическом материале сложно. Достоверно известно, однако, что потомки данной группы рептилий — птицы — имеют интенсивный обмен веществ и эндотермную систему терморегуляции (теплокровность).
Непосредственные потомки текодонтов — ранние динозавры, как ящеротазовые, так и птицетазовые — были активными хищниками, как минимум часть времени передвигавшимися на задних конечностях, что способствовало их быстрому прогрессу, особенно после вымирания большинства синапсидов на рубеже пермского и триасового периода. В юрском периоде почти все более-менее крупные наземные животные (свыше метра в длину) были динозаврами.
Постепенно часть динозавров переходит на растительную пищу, занимая освободившиеся экологические ниши. Большинство растительноядных видов вновь возвращается к передвижению на четырёх конечностях, эволюция же хищников продолжает идти по «двуногому» пути (например, у тираннозавра и тарбозавра передние конечности оказались практически редуцированны, а у манирапторов — напротив, развились в специализированную хватательную форму). В течение 140 млн лет, вплоть до конца мелового периода, основными наземными хищными позвоночными были бипедальные динозавры, некоторые формы которых по большинству признаков неотличимы от птиц.
Как видно, исторически для рептилий было свойственно главным образом именно активное передвижение на двух или четырёх выпрямленных конечностях, именно такие бегающие формы занимали господствующее положение в данном классе (парафилетической группе) животных на протяжении большей части его истории. Двуногие хищные формы изредка возникали у птиц — например фороракосы. В настоящее время подобные формы среди рептилий не возникают, поскольку экологические ниши, связанные с активным преследованием добычи, ныне заняты млекопитающими, и организм современных рептилий требует значительной перестройки для их освоения. Некоторое исключение составляет насекомоядная плащеносная ящерица, и некоторые другие агамовые, способные к бегу на двух ногах (при сохранении расположения конечностей по бокам от туловища). Такая особенность позволяет им обходить ограничение Карриера (невозможность бежать на четырёх ногах и дышать одновременно) и активно преследовать добычу или спасаться от хищников. Ящерица шлемоносный василиск во время бипедальной локомоции способна также бегать по поверхности воды. Тегу и вараны также научились обходить ограничение Карриера, но за счет модификации дыхательной системы, а не подъёма на задние конечности.
Голос |
Большинство рептилий не имеет настоящего голосового аппарата и производят лишь самые примитивные звуки вроде шипения или свиста. Крокодилы рычат, издают лающие и рокочущие звуки, детеныши квакают. Черепахи в некоторых случаях (испуг, брачный сезон, боль) могут издавать писк или стон. Представители гекконовых ящериц издают самые разные звуки — писк, визг, щебет, щелчки, крики[18].
Хозяйственное значение |
Значение пресмыкающихся для человека сравнительно невелико. Кожа крокодилов, крупных змей и ящериц применяется в кожевенной промышленности для изготовления чемоданов, ремней, обуви и т. п., однако эти предметы носят эксклюзивный характер, являясь предметом роскоши. Мясо и яйца многих черепах употребляются в пищу. Также могут употребляться в пищу некоторые ящерицы и змеи. Змеиный яд находит применение в медицине. Многие змеи полезны истреблением грызунов, а ящерицы — насекомых. Некоторые виды пресмыкающихся держат в качестве домашних животных. Аллигаторы, кайманы и некоторые крокодилы иногда разводятся на специальных фермах, для получения мяса, яиц и кожи.
Большую опасность для людей представляют ядовитые змеи, особенно в тропических странах. Многие крокодилы опасны для человека, наносят значительный ущерб животноводству. Пресноводные черепахи вредят рыбному промыслу. Крупные вараны, питоны и удавы так же могут быть опасны, будучи способными травмировать или даже убить человека.
Эволюция пресмыкающихся |
Первые представители пресмыкающихся — котилозавры — известны со среднего карбона. К концу периода появляются зверообразные, которые в пермском периоде расселяются практически по всей суше, становясь одной из господствующих групп и составляя конкуренцию пресмыкающимся. В мезозойской эре наступает расцвет пресмыкающихся, среди представителей наблюдается наибольшее многообразие. Происходит освоение морских и речных водоёмов, а также воздушного пространства. В мезозое происходит формирование всех современных групп пресмыкающихся. Последняя группа — змеи — сформировалась в меловом периоде.
В конце мелового периода происходит резкое сокращение количества видов пресмыкающихся. Однозначно указать причины вымирания современная наука пока не может.
Филогенетика |
Amniota |
| ||||||||||||||||||||||||||||||
Модельные объекты |
В 2011 году был расшифрован геном пресмыкающегося — ящерицы анолис каролинский[19]. Таким образом, это пресмыкающееся вошло в круг модельных объектов генетики.
Классификация |
В классификации пресмыкающихся много неясного, во многом потому, что большая их часть вымерла.
Класс Пресмыкающиеся (Reptilia)
Подкласс Анапсиды (Anapsida)
† ? Отряд Котилозавры (Cotylosauria)
† Отряд Captorhinida
† Отряд Мезозавры (Mesosauria = Proganosauria)
† Отряд Проколофономорфы (Procolophonomorpha)
Подкласс Диапсиды (Diapsida)
† Отряд Ареосцелидии (Araeoscelidia)
Группа Эозухии (Eosuchia)
† Группа Avicephala
Группа Neodiapsida
† Отряд Юнгинии (Younginiformes)
† ? Отряд Hupehsuchia
† ? Отряд Thalattosauriformes
† ? Надотряд Ихтиоптеригии (Ichthyopterygia)
† Отряд Ихтиозавры (Ichthyosauria)
† Хористодеры (Choristodera)
Инфракласс Лепидозавроморфы (Lepidosauromorpha)
† ? Надотряд Зауроптеригии (Sauropterygia)
† Отряд Eosauropterygia
† Отряд Нотозавры (Nothosauria)
† Отряд Плакодонты (Placodontia)
† Отряд Плезиозавры (Plesiosauria)
- Надотряд Лепидозавры (Lepidosauria)
- Отряд Клювоголовые (Rhynchocephalia)
- Отряд Чешуйчатые (Squamata): ящерицы, змеи и двуходки
Инфракласс Архозавроморфы (Archosauromorpha)
† Отряд Проторозавры (Protorosauria)
† Отряд Prolacertiformes
† Отряд Ринхозавры (Rhynchosauria)- Отряд Черепахи (Testudines или Chelonia)[20]
- Группа Архозавры (Archosauria)
- Группа Круротарзы (Crurotarsi) или Псевдозухии (Pseudosuchia)
† Отряд Фитозавры (Phytosauria)
† Отряд Rauisuchia
† Отряд Этозавры (Aetosauria)- Надотряд Крокодиломорфы (Crocodylomorpha)
- Отряд Крокодилы (Crocodilia) — единственные выжившие современные представители
† Группа Avemetatarsalia
† Клада Aphanosauria
† Группа Орнитодиры (Ornithodira)
† Отряд Птерозавры или летающие ящеры (Pterosauria)
† Группа Dinosauriformes
† Семейство Silesauridae
† Надотряд Динозавры (Dinosauria)
† Отряд Ящеротазовые (Saurichia) — вероятно дали начало птицам
† Отряд Птицетазовые (Ornithischia)
- Группа Круротарзы (Crurotarsi) или Псевдозухии (Pseudosuchia)
См. также |
- Пресмыкающиеся, описанные в 2010 году
- Пресмыкающиеся, описанные в 2009 году
Примечания |
↑ Laurin, Michel; Gauthier, Jacques A. Amniota. Tree of Life Web Project (1996). Проверено 25 января 2010. Архивировано 3 февраля 2012 года.
↑ Впервые на русском: Ренессанс динозавров — PaleoNews. paleonews.ru.
↑ Laurin, Michel; Gauthier, Jacques A. Amniota. Tree of Life Web Project (1996).
↑ The Reptile Database
↑ Высшие таксоны животных: данные о числе видов для России и всего мира
↑ См. также Список пресмыкающихся России
↑ 12 Кэрролл Р. Палеонтология и эволюция позвоночных: В 3 т. Т. 2. — М.: Мир, 1993. — 283 с. — ISBN 5-03-001819-0.
↑ Seymour, Roger S.; Smith, Sarah L; White, Craig R.; Henderson, Donald M.; Schwarz-Wings, Daniela (2012). «Blood flow to long bones indicates activity metabolism in mammals, reptiles and dinosaurs». Proceedings of the Royal Society B. 279(1728): 451—456. doi:10.1098/rspb.2011.0968. PMC 3234558 . PMID 21733896.
↑ Giant Sea Reptiles Were Warm-Blooded? (12 июня 2010). Проверено 31 мая 2018.
↑ 12 Seymour, R. S., Bennett-Stamper, C. L., Johnston, S. D., Carrier, D. R. and Grigg, G. C. (2004). «Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution». Physiological and Biochemical Zoology. 77 (6): 1051—1067. PMID 15674775. doi: 10.1086/422766.
↑ Labra A., Voje K. L., Seligmann H., Hansen T. F. (2010). “Evolution of the third eye: a phylogenetic comparative study of parietal-eye size as an ecophysiological adaptation in Liolaemus lizards”. Biological Journal of the Linnean Society. 101 (4): 870—883. DOI:10.1111/j.1095-8312.2010.01541.x..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
↑ Tosini G. (1997). “The pineal complex of reptiles: physiological and behavioral roles”. Ethology Ecology & Evolution. 9 (4): 313—333. DOI:10.1080/08927014.1997.9522875.
↑ А. Ромер, Т. Парсонс. Анатомия позвоночных. Том 1 Альфред Ромер, Томас Парсонс. скачать бесплатно. Читать он-лайн. (рус.). log-in.ru. Проверено 31 мая 2018.
↑ Анатомия позвоночных. Том 2 — Биологический кружок - Юные Исследователи Природы. uipmgu.ru. Проверено 31 мая 2018.
↑ Glenn J. Tattersall, Cleo A. C. Leite, Colin E. Sanders, Viviana Cadena, Denis V. Andrade. Seasonal reproductive endothermy in tegu lizards (англ.) // Science Advances. — 2016-01-01. — Vol. 2, iss. 1. — P. e1500951. — ISSN 2375-2548. — DOI:10.1126/sciadv.1500951.
↑ Jorge Cubo, Meike Köhler, Vivian de Buffrénil. [https://onlinelibrary.wiley.com/doi/abs/10.1111/let.12203 Bone histology of Iberosuchus macrodon
(Sebecosuchia, Crocodylomorpha)] (англ.) // Lethaia. — 2017-03-27. — Vol. 50, iss. 4. — P. 495–503. — ISSN 0024-1164. — DOI:10.1111/let.12203.
↑ Frair W, Ackman RG, Mrosovsky N (1972). «Body Temperatures of Dermochelys coriacea: Warm Turtle from Cold Water». Science. 177 (4051): 791—793. doi:10.1126/science.177.4051.791. PMID 17840128.
↑ Морозов В. П. Занимательная биоакустика. Изд. 2-е, доп., перераб. — М.: Знание, 1987. — 208 с. + 32 с. вкл. — С. 63-65
↑ Рудый, Юлия Рудый Впервые расшифрован геном пресмыкающегося. membrana.ru (05.09.2011). Архивировано 3 февраля 2012 года.
↑ Crawford, Nicholas G.; Faircloth, Brant C.; McCormack, John E.; Brumfield, Robb T.; Winker, Kevin; Glen, Travis C. (2012). «More than 1000 ultraconserved elements provide evidence that turtles are the sister group to archosaurs»(PDF). Biology Letters. 8(5): 783—786. DOI:10.1098/rsbl.2012.0331, PMC 3440978 .
Литература |
- Наумов Н. П., Карташев Н. Н. Часть 2. Пресмыкающиеся, птицы, млекопитающие // Зоология позвоночных. — М.: Высшая школа, 1979. — С. 272.
- Карташев Н. Н., Соколов В. Е., Шилов И. А. Практикум по зоологии позвоночных. — М.: Высшая школа, 1981.
- Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989.
Жизнь животных. Энциклопедия в шести томах. Том 5. (Земноводные. Пресмыкающиеся). Общая редакция члена-корреспондента АН СССР профессора Л. А. Зенкевича. — Москва: Просвещение, 1969. — 488 стр.
Ссылки |
- Пресмыкающиеся // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
Классификация современных пресмыкающихся (англ.)
Словари и энциклопедии | Большая российская · Брокгауза и Ефрона · Малый Брокгауза и Ефрона · Britannica (онлайн) |
|---|---|
| Нормативный контроль | BNE: XX526813 · BNF: 11932757v · GND: 4076664-0 · LCCN: sh85112964 · NDL: 00563034 · SUDOC: 027242544 |
Классы хордовых (Chordata) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||
| Бесчерепные (Acrania) |
| ||||||||||||
| Оболочники (Tunicata) |
| ||||||||||||
Позвоночные (Vertebrata) или Черепные (Craniata) |
| ||||||||||||
† — вымерший таксон, * — парафилетический таксон | |||||||||||||


